
ISSN (0970-2083)
ISSN (0970-2083)
Department of Biochemistry, Madurai Kamaraj University, Madurai, India
Department of Pharmaceutical Biotechnology, Ultra College Pharmacy, Madurai, India
Visit for more related articles at Journal of Industrial Pollution Control
Chromium (VI) compounds have been found to produce carcinogenic and toxic effects in animals. In rats, chromium (VI) caused more damage in liver after an i.p. administration of K2CrO4. The liver tissue accumulated with K2CrO4 and make to protein bound to DNA, nuclear protein complexes were formed in liver after treated with K2CrO4. DNA crosslinks were found in nuclei isolated from the liver of rats treated with K2CrO4. Liver nuclei contained Protein associated DNA single strand breaks in addition to DNA-Protein cross links (DPCs) complexes isolated from both the control and K2CrO4 treated cells were analyzed by gel electrophoresis. The band appeared after treatment of potassium chromate indicates that 65 kDa acidic proteins crosslinked to DNA. The agarose gel electrophoresis shown the presence of average size of DNA was approximately 24000 to 750 base pairs as compared with standard marker. Chromium (VI) has been classified as an initiator of carcinogenesis on the basis of its genetic toxicity.
Chromium (VI), K2CrO4, DPCs, Gel electrophoresis, DNA
Chromium (Cr) is ubiquitous in the environment, occurring naturally in soils, rocks, and living organisms. Cr exists in primarily two valence states, Trivalent Cr (III) and hexavalent Cr (VI), with the latter primarily produced by anthropogenic sources. Cr (III) and Cr (VI) are produced by many different industries including welding, chrome plating, chrome pigmenting, the ferrochrome industry and leather tanning ( Fishbein, 1981). It is estimated that several hundred thousand workers are potentially exposed to high level of chromium (VI) (IARC 1990).
Hexavalent chromium compounds Cr (VI) are recognized human and animal carcinogen (Mattagajasingh et al. 1996). Chromium (VI) compound administered to animal produced tumors but chromium (III) compounds do not produce them (ERA 1984). Mutagenicity of chromium (VI), but not chromium (III) has been well documented in various bacterial assay systems (Langard 1990). Interestingly, Cr (VI) does not bind to DNA or proteins in cell free system (Lay and Levina, 1998).
However, Cr (VI), which exists as an oxyanion at physiological pH, is readily transported into the cell through the cell’s sulfate anion transport system. Inside the cell Cr (VI) is believed to be reduced by the cell’s redox system to its biologically most stable form, chromium (III) (Kitagawa, 1988). Cr (III) binds to DNA as well as proteins in cell free system and has high affinity for many other biological ligands. Cr (III), However is poorly taken up into the cell and is considered to be noncarcinogenic. (Linderg and Vesterberg, 1983). During the intracellular reduction of Cr (VI) to Cr (III), reductive species such as intermediate valence states of chromium and active oxygen species are generated. Which may in turn, initiate the carcinogenic process by altering the structure of DNA.Although chromate-induced DNA protein complexes are implicated in chromate carcinogenicity, the mechanism of their formation, Composition, and biological significance are not well understood. Therefore, it is necessary to know the identity of the protein that participate in chromate- induced DNA-Protein complexes and the nature of their interaction with DNA. In the present study, we have analyzed the proteins complexed to DNA by chromate treatment of animal model. The composition and stability of chromate induced DNA-Protein complexes and the effect of antioxidants on the formation of such complexes have also been reported here.
Animal study
Male Wistar rats 100-120g were held for 5 days before experiment. Animals were divided into three groups and injected as follows: 1. Control normal saline 1ml/Kg body weight, 2. K2CrO4 was injected at a dose of 10 u mol/Kg and 3. K2CrO4 at 25u mol /Kg of body weight by introperitoneal (i,p). The rats in all groups were sacrificed 10 days post treatment. The liver was removed and washed with saline to remove all bloodstains then liver tissues were used for further analysis. The experimental animals were treated with K2CrO4 with the dose of 10, 25μ mol/ kg of body weight by intraperitonially. The control animals were injected with 0.1 mL saline.
Determination of LD50
Animals were treated with various concentration of potassium chromate (K2CrO4) ranging from 10- 100umol/Kg of body weight of the animal through intraperitoneal. A 50% reduction in mortality was observed at 50umol of K2CrO4. Therefore 10 and 25umol concentration were taken as experimental treatment for further studies on the based of subacute toxicity for intraperitonial route of potassium chromate has determined. A control animal was maintained under identical environmental condition.
Isolation of DNA-Protein complexes
The method used to isolate DNA-protein complexes was modified previously described by Mattagajasingh & Misra (1995). The rats were scarified and liver removed after 10 days of experiments. Potassium chromate treated and control animal liver cells were collected by centrifugation at 10,000 rpm for 10 min and were washed three times in phosphate buffered saline. The cell were lysed in 10 mL of 10 mM Tris containing 2% SDS, 1mM HEPES/PMSF (pH 7.5) by shaking on a platform shaker for 2h at room temperature. The cell lysates were transferred into tight fitting homogenizer and given 10 min strokes.
The samples were sedimented at 10,000 rpm for 30 min at -4°C, using refrigerates centrifuge. The pellets were placed in a solution5mL of 5M urea containing 1mM PMSF and rocked on platform shaker at -40C for 30 min. The samples were again homogenized as above, and then SDS was added to 2% final concentration. The DNA-protein complexes were isolated by sucrose centrifugation and were rinsed in 10 mM Tris-HCl (pH 7.5) containing 2% SDS. The final pellets were suspended in 10 mM Tris-HCl containing 1mM PMSF (pH 7.5) in eppendroff tubes by gentle pippeting and overnight incubation at -200C.
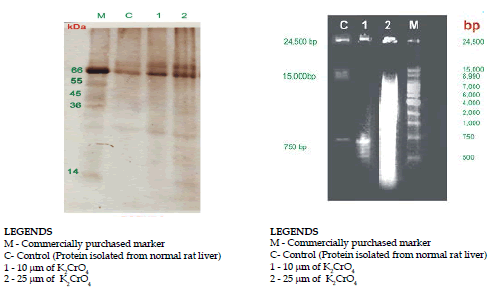
The DNA-Protein complexes were precipitated in 70% acetone at -20°C. The DNA protein complexes were collected by centrifugation at 10,000 rpm for 15 min at -4°C, pellets were rinsed in 70% acetone, and resuspended in 1 ml of 10mM Tris-HCl containing 1 mM PMSF (pH 7.5) by gentle pippeting or by shaking on shaker for 15 min at -40C. The DNA content and purity of the sample were determined by measuring the absorbance at 260 and 280 nm. The protein concentrations cell lysates were determined by using Lowry’s method (1951) and BSA as the standard. The protein complexes separated by 15% SDS PAGE, analytical electrophoresis of proteins was carried out under conditions that ensure dissociation of the proteins into their peptide subunits and that minimize aggregation. The protein complezes were resolved in SDS-PAGE by continuous buffer system (Laemmli 1970).The sample volume containing 150-200μg proteins were loaded in the wells and electrophoresis was carried out at a constant voltage of 100 V across the gel. The gel stained with silver nitrate and the protein band has visualized by gel documentation system.
Nuclei purified from DNA-Protein complexes
Nuclei were sedimented from homogenization of liver tissue at 7000 rpm for 15 min at -4°C resuspended in 10 mM Tris-Hcl containing 250 mM nuclease free sucrose, 3mM MgCl2 and 1mM PMSF (pH 7.5); and layered over a similar solution but containing 880 mM sucrose. Nuclei were subsequently collected by centrifugation for 10 min at 10,000 at -4°C using refrigerated centrifuge. The extracts of purified nuclei DNA were applied to horizontal agarose gel electrophoresis (1.0%) prepared in Tris Borate buffer. The gel run was carried out at 20v 120min, Gel was stained with ethidium bromide(1μg/mL) and the bands were visualized under UV illumination and the results were photographed (Sambrook et al. 1989).
The LD50 determination results of potassium chromate for intraperitoneal route indicated that 50% mortality was observed at 60μmol of potassium chromate. (As shown in Fig.1) Therefore 10, and 25 μmol concentration of potassium chromate were taken as experimental treatment. The animals were scarified after 10 days and the liver tissue washed with saline to remove blood. The DNA protein complexes were prepared from the liver tissue and used for the analysis; there was increased complexes formation on the basis of dose dependence. These resulted in dose dependent increases in the formation of DNA-protein complexes in liver tissues. Whenever increasing the concentration of K2CrO4 treatment resulted increasing DNA-protein complexes as compared with the control. K2CrO4 induced DNA-protein complexes formation in liver was also found to be time dependent a shown in Fig.2.

Fig. 1 Determination of LD50

Fig. 2 Dose dependent in the formation of DNA-Protein complexes
The DNA-protein complexes formation increase in a time dependent time manner in a animal treated with 25μmol K2CrO4 for different time period and after 10 days a 15 fold increases in the formation of DNA-protein complexes was observed as compares with the control tissues.
Analysis of Protein Complexes by PAGE
DNA-Protein complexes isolated from both the control and K2CrO4 treated cells were analyzed by gel electrophoresis. DNA-Protein complexes were loaded on the acidic end of the gel in order to avoid the entry of nucleic acids into the gel. Silver staining of the gel of DNA-Protein complexes isolated from control animal. Protein complexes isolated from liver tissue were separately by 1.5% SDS PAGE.
Figure 3 has shown the presence of highly complexes proteins in the SDS PAGE silver staining. The proteins that were complexed to DNA, in cells exposed to 25μmol K2CrO4 were r extraction as shown in Fig 3. Analysis of the molecular weight of these proteins showed that the protein has molecular mass of 65 kDa. The proteins cross-linked to DNA upon K2CrO4 exposure was found to be dependent on the dose of K2CrO4 because animal treated with 25μmol K2CrO4 after 10 days had many other proteins in addition to the above proteins complexed to DNA. Since 25umol K2CrO4 treated cells were found to be killed by such treatments (25μmol), there was a reason to believe that the additional proteins cross-linked to DNA could be due to dead cells. This treatment did not affect cell viability but cross-linked the same proteins to DNA.
There is of course, a possibility that the cytoplasmic proteins might associate with DNA during the cell lyses. We analyzed DNA-Protein complexes isolated from either whole cells or purified intact nuclei of cells treated with KK2CrO4 . That was not the case in illustrated by the fact that identical proteins were found to be complexed to DNA when either whole cells or nuclei of cells treated with K2CrO4 were used as starting material. Hence, it appears likely that K2CrO4 induces the cross-linking of nuclear proteins to DNA.
As shown in this Figure 3, all of the proteins cross-linked to DNA by 25μmol K2CrO4 treatment and a 65 kDa acidic protein cross-linked to DNA by higher doses of K2CrO4 were found in this fraction. These results suggest that nuclear matrix proteins are the targets for chromate-induced DNA-Protein cross -linking. The chromate-induced DNA crosslinks, which were DNA-Protein in nature, effectively masked the single strands breaks. Liver nuclei from chromate-treated rats caused DNA elution rates to increase slightly compared to control.
Stability of DNA Protein complexes/ DNA agarose gel
The stability of DNA-Protein complexes was tested by monitoring the recovery of DNA and protein in the following treatment of DNase I, RNase and proteinase K, ß-mercaptoethanol. The average size of DNA was approximately 24500, 15000 and750bp pairs bands observed by gel. (Fig.4). The control sample had almost 100% recovery of both DNA and protein in the pellet by centrifugation. A treatment of DNA protein complexes did not interfere with recovery of DNA or protein. Proteinase k treatment dissociates most of protein from the DNA-Protein complexes without affecting the recovery of DNA.
In the present study, Chromium (VI) caused more damage in rat liver after an i.p administration of K2CrO4. The level of chromium accumulated in rat liver has been measured after 10 days, of treatment with K2CrO4 . The liver tissue was accumulated large of chromium and concentrated in the nucleus. Chromium was bound to DNA, nuclear protein complexes were formed in liver . The significant amounts of chromium were bound to DNA and non histone proteins. Chromium enters the liver at faster way and increasing the level of chromium bound to liver nuclear protein was observed in the experimental samples. It is possible that Cr (VI) induces a cytoplasmic factors, which influences the uptake and subsequent binding to nuclear protein complexes. Chromium DNA adducts was formed after K2CrO4 on the oxidation state of the chromium compound administered. Our data suggested that certain type of Cr-DNA complexes form after administration of Cr (VI) not produce lesion on the DNA. Liver nuclei contained protein associated DNA single strand breaks in addition to DNA-protein cross-links. The DNA damage induced by chromate is discussed in relation to the carcinogenicity and toxicity of chromium (VI) compounds.
Chromium (VI) containing compounds are recognized occupational carcinogens and pose an environmental health risk. Chromium (VI) has been classified as an initiator of carcinogenesis on the basis of its genetic toxicity.
This research was supported by the grant from Human resource Development group, CSIR, New Delhi
Copyright © 2025 Research and Reviews, All Rights Reserved